- 中国科学论文统计源期刊
- 中国科技核心期刊
- 美国化学文摘(CA)来源期刊
- 日本科学技术振兴机构数据库(JST)
红细胞与血小板储存损伤新机制及其应用研究
中国输血协会临床输血管理学专业委员会组稿
深圳市第二人民医院输血科组织编译
引 言
本期《输血医学新进展》聚焦红细胞与血小板储存损伤新机制及其应用,通过分享《Blood》杂志发表的3篇文章,展示了该领域的一些最新研究成果,内容涵盖铁死亡调控储存的小鼠和人红细胞的溶血过程,供血者遗传与生物学特征对犬尿氨酸代谢的调控影响红细胞在体内外的溶血,以及抑制RHOA活性可维持长期低温保存血小板的存活和止血功能。这些研究成果具有重要意义:1)对红细胞储存质量相关等位基因进行遗传筛查,为新型血液管理策略提供依据,通过基于遗传学预测的储存稳定性来定制浓缩红细胞的保存期限;2)犬尿氨酸(色氨酸代谢产物)是与储存红细胞渗透脆性相关的新型代谢和遗传标志物,其水平受处理策略、供者特征及遗传背景影响;首次通过mQTL 研究揭示犬尿氨酸通路限速酶编码区多态性与储存 RBC 在体内外溶血倾向的关联性;3)研究证实RHOA激活在血小板短期与长期低温保存损伤中的关键作用,通过使用R-G04或S-G04抑制RHOA,可预防低温保存引发的血小板储存损伤,为延长血小板保存期限提供了有效解决方案。这些新的研究成果为优化红细胞、血小板的储存与管理策略提供了新的理论依据。
铁死亡调控储存的小鼠和人红细胞的溶血过程
编译:于姣姣,胡晓榕 审校:伍昌林
1研究背景
红细胞储存期间会积累蛋白质和脂质的氧化损伤,这是一种逐渐不可逆的现象。“储存损伤”促进了受损成分的囊泡化,导致形态不规则、较小的红细胞,这些红细胞更容易发生袋内溶血、血液循环中的血管内溶血或通过脾脏和/或肝脏隔离和红细胞吞噬作用导致的血管外溶血。红细胞储存质量的黄金标准通过两项指标判定:袋内溶血率(<1%)和输注后回收率(PTR>75%)。对新鲜和储存的红细胞代谢和溶血异质性的遗传基础的研究为了解驱动红细胞对氧化剂、溶血性损伤的反应机制提供了一个窗口。最近研发的杰克逊实验室杂交小鼠(J:DO)模型包含比传统杂交小鼠更高水平的遗传多样性,该资源群体能够以与基于人群的研究相同的方式捕捉遗传异质性的影响,同时仍保留小鼠实验优势,包括相关因素的标准化以及对环境变异来源的严格控制,从而为研究遗传变异如何影响红细胞代谢及储存损伤异质性提供新一代研究基础。
2主要结果
2.1 将Steap3映射为J:DO小鼠红细胞输注后恢复的关键遗传驱动因素
选择350只正在进行J:DO育种计划的第34代小鼠(图1A),这些小鼠的红细胞模拟人类血库中红细胞储存条件储存7天,储存的红细胞与新鲜的含示踪剂红细胞混合,并输注到绿色荧光蛋白阳性的小鼠中。通过流式细胞术确定PTR:输血后24小时获取的血液中,检测输血后恢复率PTR(图1A)。尽管没有捕获这个J:DO队列的完整血细胞计数,但任何小鼠都没有出现血液学异常(Hb<13.6g/dL)。此外,贫血并不是多样性育种的一种紧急属性,因为其他关于J:DO种群的研究显示,J:DO小鼠的血液中血红蛋白水平正常。小鼠在~143 000阵列上进行了基因型分析,并使用PTR作为定量性状(图1B)进行了QTL分析,识别出一条强QTL在染色体1编码的铁还原酶Steap3(图1C-D)附近。
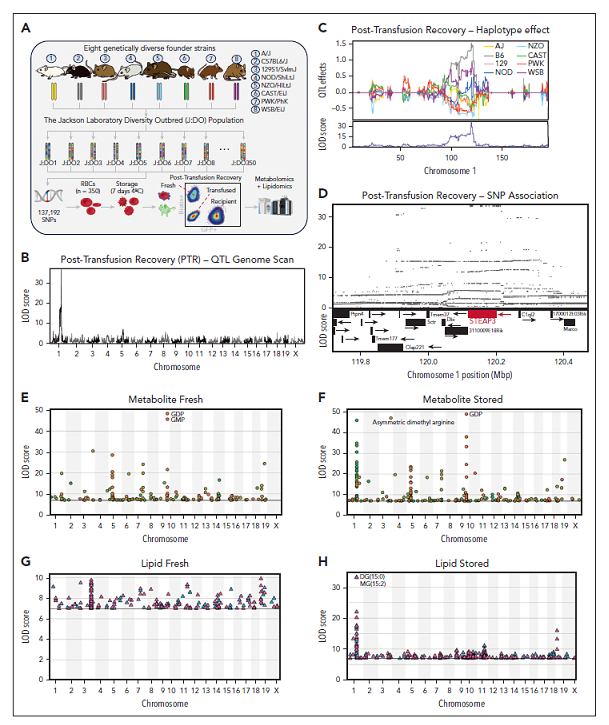
图1. J:DO小鼠红细胞代谢和储存质量异质性的遗传基础
2.2 J:DO小鼠RBC代谢异质性的遗传驱动因素
代谢组学和脂质组学数据进行遗传定位分析,识别新鲜和储存的红细胞(RBC)的QTL(代谢物mQTL和脂质lQTL)。基于严格的LOD分数>8的阈值,研究者在新鲜样本中定位76个mQTL和54个lQTL,在储存样本中定位114个mQTL和168个lQTL。研究者在染色体1、5、7、9、12和14上观察到mQTL热点;对于lQTL,在染色体3上观察到新鲜特异性热点,在染色体1、7、9、10、11、13和19上观察到储存特异性热点。研究者还推断SNP和结构性变异(SV)的基因型,并在QTL支持区域内进行关联分析,发现了一个与胸腺嘧啶水平关联更强的SV,表明它可能是实际的因果变异,胸腺嘧啶相关的SV位于胸腺嘧啶磷酸酶(Tymp)基因的上游。QTL单倍型与另一个J:DO队列中观察到的肝脏组织中Tymp的顺式eQTL高度一致,表明在Tymp具有B6和PWK等位基因的J:DO小鼠中,Tymp蛋白水平较低,导致胸腺嘧啶水平较高。
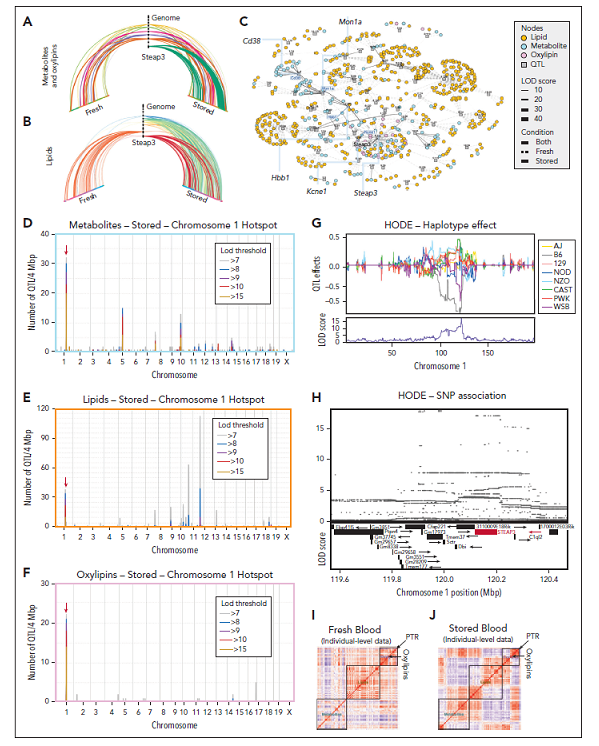
图2. 染色体l QTL热点揭示Steap3是储存RBCs中代谢物、氧脂和脂质的关键调节因子
2.3 Steap3的遗传变异与输血后回收和脂质过氧化有关
PTR的顶级负代谢相关物是氧脂类,包括前列腺素、二十碳烷和十八碳二烯酸羟基和羟基过氧化物(分别为HETEs、HODEs和HPETEs,图1K-M)。相反,链多不饱和脂肪酸是正相关物,表明这些底物的储存诱导氧化减少输血后性能下降具有保护作用(图1K)。在代谢物和脂质中寻找发现了一个广泛的QTL热点,该热点覆盖染色体1上的Steap3编码区(图2A-B)。使用QTL结果定义一个基因-代谢物对的共定位网络视图,该视图展示了STEAP3 QTL热点的中心性(图2C),如图2D-F所示。该QTL区域的单倍型效应复杂,B6和WSB等位基因与其他等位基因有明显的区别(例如HODEs,图2G)。Steap3位于QTL区域的中心,B6和WSB共享SNP变异最好地捕捉了单倍型关联(图2H)。遗传效应高度一致,并且在这个QTL热点中特异性地针对储存的RBC,这从代谢物、脂质、氧脂类和PTR之间的关系如何分解成2个相关和反相关的组中可以看出,表明它们是由STEAP3的遗传变异驱动的(图2I-K)。
2.4 代谢组学和脂质组学QTL资源网络工具
研究者开发了一个公开在线的门户网站,使用QTLViewer网络工具进行QTL结果和处理数据分布探索(https://churchilllab.jax.org/qtlviewer/Zimring/RBC),功能包括从PTR代谢物以及脂质表型中绘制和导出结果,包括检查感兴趣的协变量与性状、QTL扫描、SNP关联单倍型效应扫描的相关性的能力,还确定了其他一些QTL热点处的强候选基因,这些基因在图2A-C中以蜂窝图和网络可视化。除第1号染色体上的热点外,研究者还确定了其他代谢物-基因关联。
2.5对STEAP3的基因操作调节储存小鼠红细胞中的脂质过氧化
研究者首先将来自FVB/J小鼠的高表型STEAP3引入C5BL/6J背景,这增加了脂质过氧化和降低了PTR(约25%)。在储存良好的小鼠中,研究STEAP3基因敲除(KO)小鼠,这也对储存中的红细胞代谢产生显著影响(图2L)。具体来说,Steap3 KO和功能获得(GOF)都导致小鼠PTR的降低(约45%),Steap3 KO与GOF小鼠相比仍然2倍(图2M)。然而,只有GOF小鼠,而不是KO小鼠,在储存结束时的红细胞中显示出提高的氧化脂水平(图2N)。
2.6氧脂素、铁和STEAP3与13 000名REDS RBC omics供体的溶血有关
研究者首先确定REDS RBC Omics研究中13,091个索引和643个供体(图3A)的溶血测定的代谢基础。研究发现氧脂是索引和召回血单元储存溶血和囊泡化的主要相关因素(图3B-C)。在REDS队列中监测了37个STEAP3 SNP:23位于内含子中,3个位于下游3'-非翻译区,而11个位于编码区,其中2个为同义词,9个为非同义词(图3C)。在储存的人类RBC中,STEAP3变异与溶血(图3C),这些常见变异位于Steap3酶活性位点附近的区域(图3D)。Steap3 SNPs和脂质过氧化的等位基因频率在人口数据的二维均匀流形近似和投影表示重叠(图3E)。
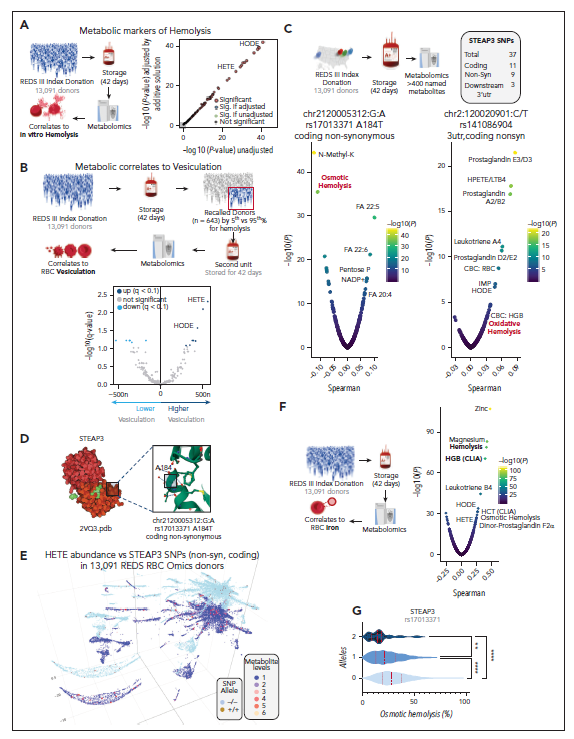
图3. 氧脂素、铁和STEAP3与13000名REDS RBC Omics献血者的溶血有关
2.7遗传因素导致13 091个储存的人红细胞中异质性脂质过氧化
在索引和召回的供体中,携带1或2个非同义编码SNPs的STEAP3等位基因与渗透和储存溶血(图3G-H)、红细胞血红蛋白和铁含量以及G2(图4A-K)呈负相关。氧化脂质基因突变的组合阐明由L-肉碱、NADPH和谷胱甘肽依赖系统驱动的脂质解毒途径(图4I),该图与铁死亡领域的文献有重叠。在所有氧脂素中,HETEs(图4B)显示出最高数量的命中,这些大部分与其他氧脂素共享,包括HPETE、前列腺素A2和D2/E2、白三烯A4(图4)和HODEs。此外,环氧水解酶2成为二硝酸前列腺素F2α(图4F)和前列腺素G2(图4H)最显著命中。与Steap3 SNPs类似,LPCAT3、FADS1/2和EPHX2的等位基因频率与更高的(EPHX2和LPCAT3)和更低的(FADS1/2)脂质过氧化和渗透压损伤时的溶血倾向显著相关(图5A-I);种族划分的等位基因频率显示,在非洲血统的献血者中,LPCAT3和EPHX2显著富集,而FADS1/2则代表性不足(图5J)。

图4. 人类红细胞在储存42天后,异质性脂质过氧化的遗传因素
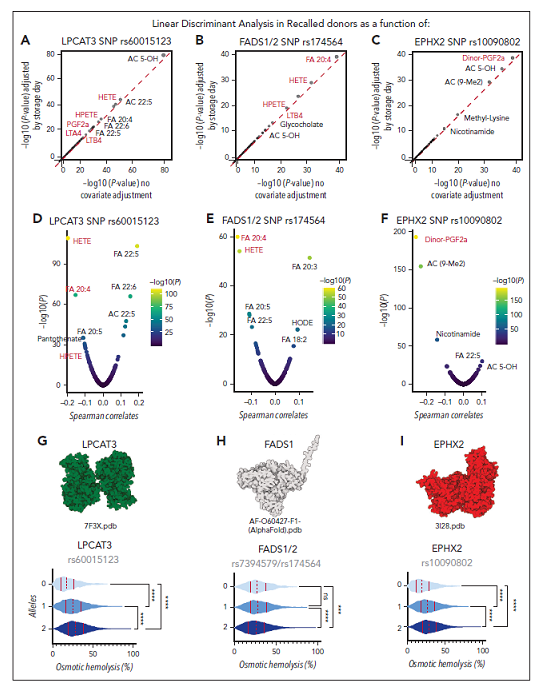
图5. LPCAT3、FADS1/2和EHPHX2的遗传多态性
2.8 TP53在健康献血者中是多态性的,并与脂质过氧化和溶血的遗传调节因子有关
研究者对REDS红细胞组学项目中13,091名献血者的50多个常见TP53单核苷酸多态性(SNP)进行了评估(图6A展示其中最常见的20种等位基因)。该列表包含与癌症易感性增加相关的常见SNP(图6A)。值得注意的是,非编码SNP rs8064946的等位基因拷贝数与渗透性溶血相关,并与EPHX2、LPCAT3、G6PD、Steap3、GPX4、ACKR1和SLC22A16的等位基因频率相关(图6B)。在初始供体和回忆供体中(图6C-D),rs8064946与渗透脆性标志物及氧化还原稳态标志物(磷酸戊糖异构体、NADP+)相关。等位基因频率随种族分布的变化显示,潜在致病性rs1042522和rs105200764在非洲裔供体中患病率较低,而非编码rs8064946则患病率较高(图6E-H),且均与更高的渗透脆性相关(图6I)。仅有3名献血者携带致病性TP53 SNP rs1800371和rs1800372等位基因纯合子(TP53 DNA结合位点的H178R和R248Q突变[图6I]),而杂合基因型更为常见且与更高渗透性溶血相关。
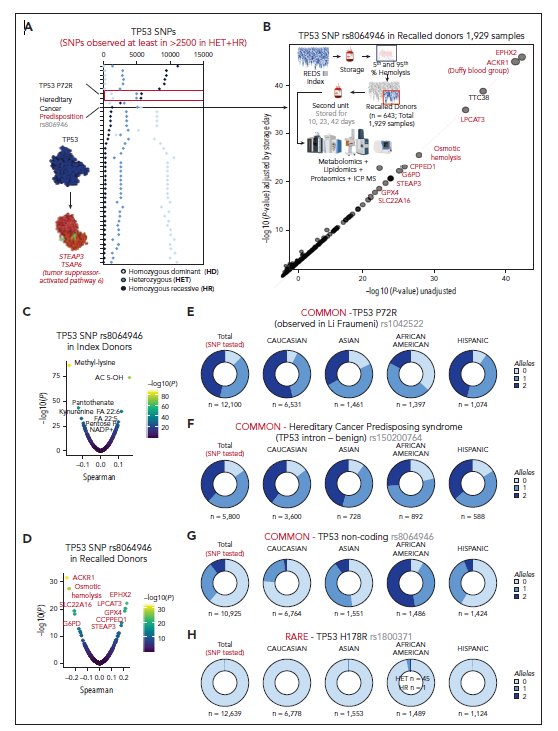
图6. TP53在健康献血者群体中是多态的,并且与溶血有关
2.9氧脂蛋白和相关的遗传特征与受体输血效率较低有关
研究者调取REDS项目的“vein-to-vein”数据库,将供体基因型或氧化脂蛋白检测数据与数千例接受REDS供体单位输血的单剂量受血者血红蛋白(Hb)增量相关联。遗憾的是,由于数据库中记录的携带相关STEAP3等位基因供体单位输血事件数量不足,无法进行这些具有转化学意义的分析。但观察发现,携带rs8064946 TP53单核苷酸多态性(SNP)双拷贝的供体单位输血,其Hb增量显著降低(P<0.001),输注单位年龄超过5周更为明显(图7A-B)。类似现象也出现在携带LPCAT3 rs60015123双拷贝的供体单位输血中,无论是输注后即刻还是3周或36-42天后(图7C-E),其Hb增量均显著下降。最后,研究发现储存结束时氧化脂蛋白水平升高的供体单位输血,其受血者的Hb增量显著降低(图7F)。
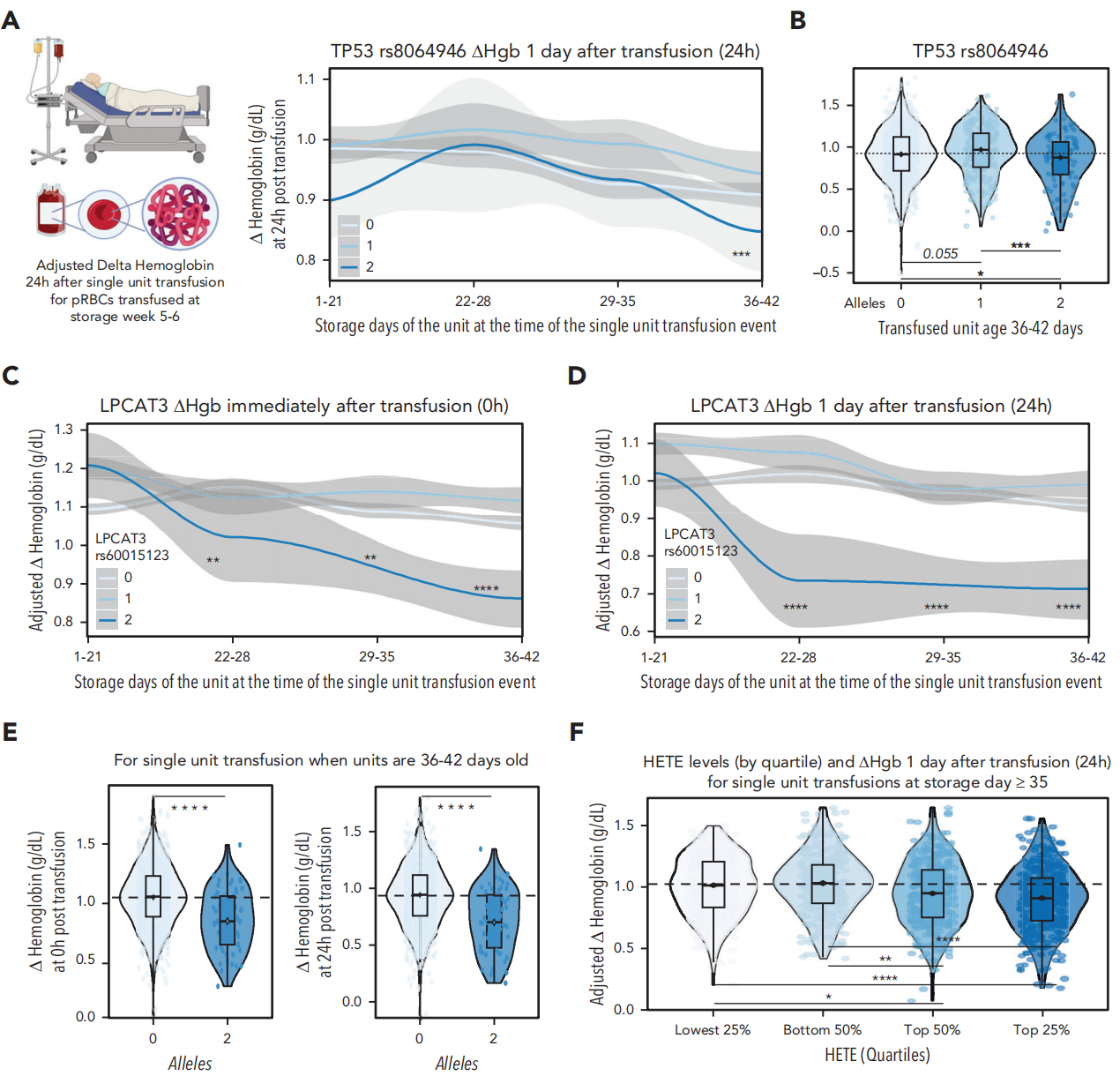
图7. 氧脂素及其调控基因的多态性与接受非自体输血的重症患者体内溶血有关
3 主要结论
1)研究发现编码Steap3蛋白的区域单核苷酸多态性(SNP)是导致储存小鼠红细胞脂质过氧化及PTR水平异常的主要驱动因素。在小鼠和人类中,STEAP3通过内体运输和囊泡化外排运输在铁代谢调控中发挥核心作用,STEAP3基因缺陷与人类贫血相关。
2)该成果可指导对与储存质量相关等位基因进行遗传筛查,为血液管理策略提供依据,通过基于遗传学预测的储存稳定性来定制浓缩红细胞的保存期限。
文献来源:Angelo D'Alessandro, Gregory R Keele , Ariel Hay , et al. Ferroptosis regulates hemolysis in stored murine and human red blood cells. Blood. 2025;145(7):765-783. doi: 10.1182/blood.2024026109.
供血者遗传与生物学特征对犬尿氨酸代谢的调控
影响红细胞在体内外的溶血
编译:叶至顺,廖奇峰 审校:伍昌林
1研究背景
血库冷藏储存条件会引发浓缩红细胞(pRBCs)一系列生化和形态学改变,统称为“储存损伤”。这些改变最终影响RBC在受者体内的循环能力,从而损害氧气的运输和递送,并对受血者产生免疫调节作用。近年来,输血医学中的组学研究加深了对pRBC储存损伤的理解,pRBC的代谢受到储存时间、处理策略以及供者暴露因素的影响。此外,供者遗传背景和其他生物学因素也影响pRBC储存和输血效果。尽管在对pRBC质量的表征方面已取得显著进展,但大多数组学研究规模有限,未能充分揭示供者生物学、处理策略或遗传背景对储存单位“代谢年龄”的影响程度,也未明确供者遗传背景是否或多大程度上影响RBC的溶血倾向和输血后疗效。本研究利用REDS-III和REDS-IV-P项目,旨在提高血液供者安全性并优化输血效果。
2主要结果
2.1 RBC代谢受储存时间及添加剂的影响
对643名回访供者队列的pRBCs分别在储存第10、23及42天进行代谢组学分析(图1A)。多变量分析显示(图1B-D),1,929份样本依据储存时间及储存用添加剂溶液形成聚类,两因素各自解释约10%的总体代谢变异。储存显著影响既往已确定的储存损伤代谢标志物,包括葡萄糖、2,3-二磷酸甘油酸及ATP的耗竭,以及乳酸、5-氧脯氨酸和次黄嘌呤的累积(图1E)。进一步分析证实储存时间及添加剂溶液为主要聚类驱动因素:尽管储存时间相近,仍识别出3个独立聚类,主要由不同添加剂解释(图1F-G)。这些组间的主要代谢差异物为添加剂组分—甘露醇和柠檬酸盐(图1H)。
2.2储存损伤的代谢标志物与REDS RBC Omics回访供者队列的溶血倾向相关性较差
首先,使用储存溶血、氧化溶血和渗透溶血参数对回访供者样本进行颜色编码(图2A-C)。尽管储存溶血和氧化溶血随储存时间延长呈增加趋势,但渗透溶血未观察到显著的储存时间依赖性。代谢物水平与溶血参数的相关性分析显示,在所有既往定义的储存损伤代谢标志物中,仅次黄嘌呤增加和腺嘌呤耗竭分别与氧化溶血和储存溶血显著相关,其余代谢标志物均未显示显著关联(图2D-F)。尽管所有供者在时间进程中均呈现显著且相似的趋势,但在最早时间点(第10天)即处于某一代谢标志物第5或第95百分位的供者,在后续所有3个时间点(储存第10、23及42天)均维持该排名(图2G)。换言之,对于某一特定代谢物(如次黄嘌呤),处于低丰度第5百分位供者的储存末期水平低于处于高丰度第5百分位供者早期储存时的水平(图2G)。

图1 .REDS RBC Omics回访供者群体的代谢组学分析
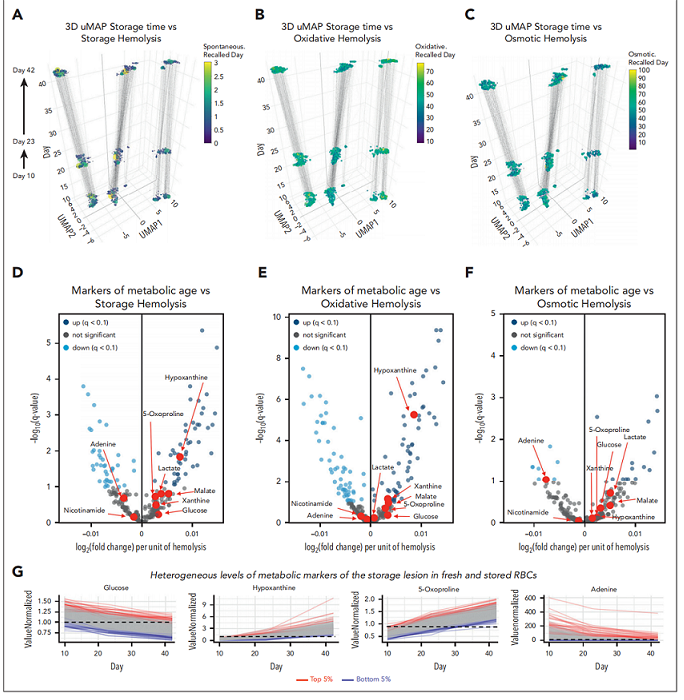
图2. 在REDS RBC Omics回访供者队列中,储存损伤的代谢标志物与溶血倾向相关性差
2.3犬尿氨酸是储存红细胞渗透脆性的标志物,受供者年龄、BMI及性别影响
两种方法均独立地将犬尿氨酸鉴定为与渗透脆性及溶血呈正相关的最显著代谢标志物(图3A-C)。展示了机器学习方法的结果及回归算法的验证测试。犬尿氨酸亦与氧化溶血呈正相关,但与储存溶血无显著关联。当分别聚焦于储存第 10、23 或 42 天时,犬尿氨酸与渗透脆性的正相关关系依然成立。我们前期对回访供者队列中 599 份样本的试点研究进行的荟萃分析亦证实,犬尿氨酸水平与渗透脆性呈正相关,尤其适用于 AS-3储存的样本(465份样本)。除处理因素外,在全部 1,929 份回访供者样本中,犬尿氨酸水平显著受供者生物学特征影响,与供者年龄及 BMI呈强正相关(图3D-H)。男性供者犬尿氨酸水平显著高于女性供者(图3I)。
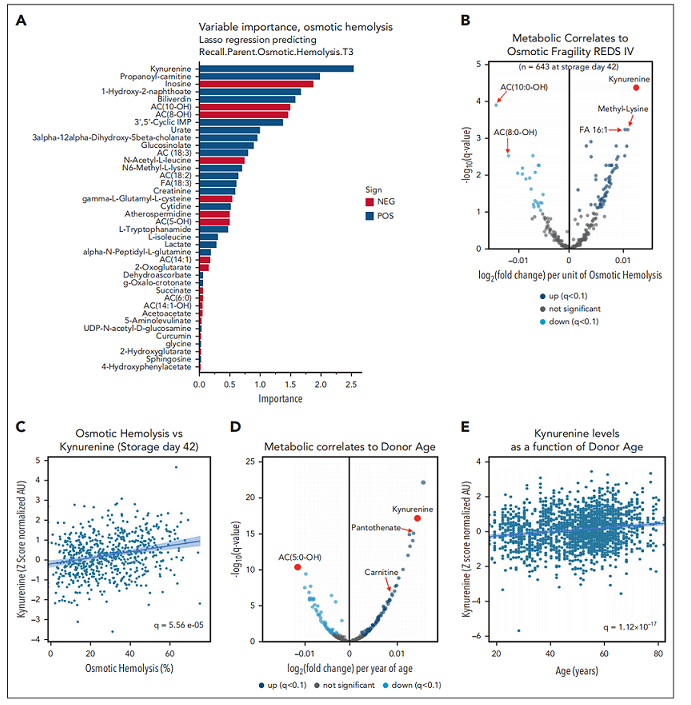
图3. 犬尿氨酸是储存红细胞渗透脆性的标志物,并受供者年龄、BMI 及性别影响
2.4回访供者样本的多组学分析揭示了犬尿氨酸升高供者红细胞渗透脆性的分子机制
针对643名回访供者单位(储存第10、23、42天),生成蛋白质组学、代谢组学和脂质组学数据,以鉴定与犬尿氨酸水平相关的组学特征(图4A)。结果显示,犬尿氨酸水平与补体成分以及纤维蛋白原α链和γ链呈正相关,同时与红细胞结构及功能蛋白水平下降呈平行关系(图4B)。在代谢水平上,犬尿氨酸升高伴随全部胞内氨基酸,尤其是芳香族氨基酸水平升高(图4C)。渗透脆性位于第5和第95百分位的供者呈现出不同的代谢表型,当犬尿氨酸水平最高与最低供者时,这一差异得以重现。与图4中的相关性分析一致,犬尿氨酸和渗透溶血水平升高均与多种氨基酸及L-肉碱和多种酰基肉碱水平升高相关,后者是膜脂修复的标志物,尤其是羟基酰基肉碱(10-OH、12-OH、14-OH和15-OH)。对回访供者队列1,929份RBC样本的多组学整合通路分析显示,参与病毒应答、补体与凝血以及氨基酸转运与代谢的组分显著富集(图4D)。

图4. REDS RBC Omics回访供者队列中与犬尿氨酸水平相关的蛋白和代谢物特征
2.5同一供者在多次献血中犬尿氨酸水平具有高度重复性
研究者在13,091名索引供者样本中验证发现(图5A),与回访供者队列类似,索引供者队列的犬尿氨酸水平呈右偏分布,部分供者的水平可达中位值的7倍(图5B)。对13,091名索引供者进行无监督uMAP聚类,结果显示犬尿氨酸水平在聚类图核心区域的特定供者亚群中显著升高(图5C)。按供者年龄绘制的犬尿氨酸水平折线图再次证实,供者年龄与犬尿氨酸水平呈正相关(图5D)。与女性供者相比,男性供者在约55岁之前均表现为更高的犬尿氨酸水平,该年龄接近美国女性绝经的平均年龄(51岁)(图5D)。供者BMI与犬尿氨酸水平亦呈正相关,从而在这一扩大20倍的验证队列中确认了回访供者队列中的发现(图3)。与渗透脆性的相关性分析显示,索引供者队列中犬尿氨酸水平与渗透脆性呈强正相关(图5E)。由于643名供者同时参与了索引和回访队列,我们比较这些供者在两次独立献血(间隔2-12个月)的储存第42天RBC中犬尿氨酸水平的相关性,以评估该代谢物在储存末期水平在同一供者间的可重复性(图5F),结果显示二者显著相关(图5F)。
2.6犬尿氨酸水平的异质性至少部分由遗传因素解释
研究者评估遗传背景对该代谢物遗传力的影响(图6A),单卵双胞胎间呈显著相关,而双卵双胞胎间无显著相关(图6B-C)。在13,091名索引供者中开展犬尿氨酸mQTL分析,该队列已通过精准输血医学芯片对879,000个SNP位点进行基因型检测。对643名回访供者中250人的试点mQTL分析因样本量不足,未鉴定出与RBC单位犬尿氨酸水平异质性相关的SNP。在更大规模的REDS RBC Omics索引供者队列中开展的mQTL分析获得了显著关联,其中效应量最大的位点位于犬尿氨酸及其前体色氨酸的转运体编码区SLC7A5(又称LAT1)(图6D),其次为ATXN2。随后的显著SNP位于犬尿氨酸合成与代谢的限速酶编码区:犬尿氨酸单氧化酶、吲哚胺2,3-双加氧酶、色氨酸双加氧酶以及另一色氨酸转运体SLC22A8(图6D)。用13C1?N-色氨酸孵育RBC,以验证成熟RBC能否摄取标记色氨酸并将其转化为犬尿氨酸。结果证实在37°C孵育16小时后,成熟RBC可摄取标记色氨酸,但其转化为犬尿氨酸的比例极低(<5%)。
2.7 SLC7A5 和 ATXN2 的多态性分别与高、低犬尿氨酸水平相关
按犬尿氨酸水平对索引供者进行颜色编码的 uMAP 显示,水平分布与 SLC7A5最强关联 SNP(rs8052118,内含子区)的基因剂量高度重叠(图6E)。犬尿氨酸是在该 SNP 纯合隐性基因型供者中水平最高的代谢物,而在显性基因型纯合供者中水平最低(图 6F)。利用芯片定量数据证实犬尿氨酸及其同通路代谢物羟吲哚乙酸分别位列与 SLC7A5 和 ATXN2 SNP 正、负相关性第一和第二强的代谢物(图6G-H)。这两个SNP 对犬尿氨酸水平呈叠加性效应:同时携带 SLC7A5 纯合隐性基因型和 ATXN2 纯合显性基因型的供者,其犬尿氨酸水平最高(图6I)。
2.8 犬尿氨酸水平及 SLC7A5 SNP 与储存红细胞在小鼠模型和临床患者输注后体内表现不佳相关
为验证储存RBC中的犬尿氨酸水平不仅与体外渗透脆性相关,还与输注后体内血管外溶血增加相关,在遗传多样性极高的小鼠群体中开展了储存及PTR(输注后回收)实验(图7A)。这些大规模小鼠研究结果显示,犬尿氨酸水平与输注后 24 小时回收率呈显著负相关(P< 0.001)。为评估这些发现的人体转化意义,利用 RBC Omics “静脉-静脉”数据库,将 SLC7A5 rs8052118 SNP 剂量与4,470名接受 REDS RBC Omics 索引供者血液制品单位输血患者的血红蛋白增量和胆红素增量关联(图 7B)。结果提示溶血发生:与携带SLC7A5 rs8052118 纯合隐性基因型供者血液制品输注的患者相比,携带纯合显性基因型供者血液制品输注的患者血红蛋白增量显著降低(图7B-C),胆红素水平显著升高(图7D)。

图5. 在REDS RBC Omics索引供者队列13,091名供者中测定储存末期的犬尿氨酸水平
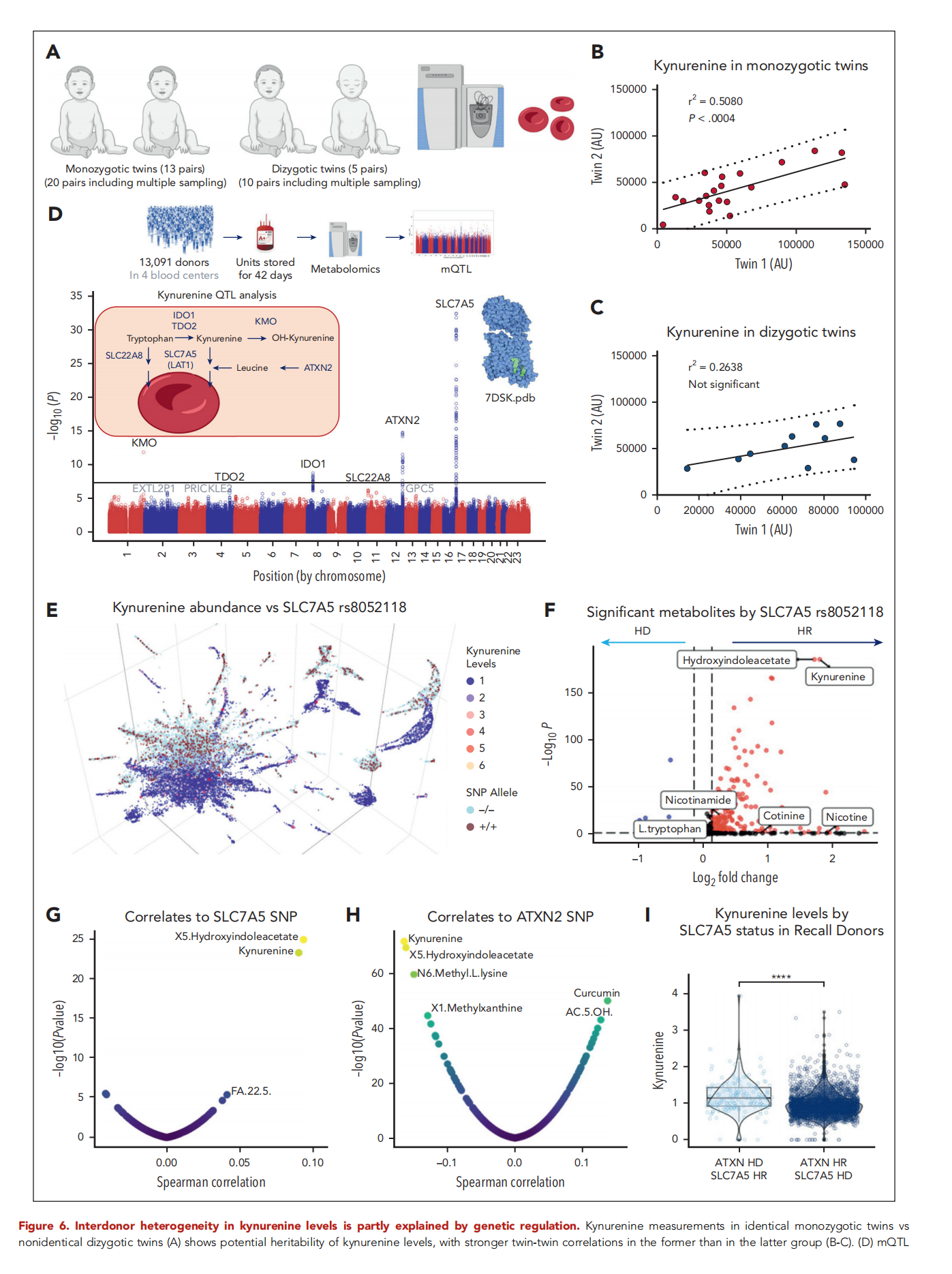
图6. 供者间犬尿氨酸水平的异质性部分由遗传调控解释

图7. 高犬尿氨酸水平及其相关遗传多态性导致小鼠储存模型及临床患者输注后血管外溶血增加,并降低血红蛋白增量
3 主要结论
1)色氨酸代谢产物犬尿氨酸是储存红细胞渗透脆性相关的新型代谢和遗传标志物,其水平受处理策略、供者特征及遗传背景影响。
2)最显著的遗传位点(SLC7A5)不仅验证近期在其他人群中开展的 mQTL 研究结果,还进一步扩展了其在体外和体内溶血方面的临床转化意义,涉及小鼠模型及数千名接受携带该SNP 的志愿供者血液制品的输血受者。
3)首次通过 mQTL 研究揭示犬尿氨酸通路限速酶编码区多态性与储存RBC体外和体内溶血倾向的关联,储存损伤的代谢标志物确实在供者间呈现一致变化趋势,但储存期间代谢物消耗或积累的速度和程度取决于献血早期这些代谢物水平的供者间异质性。
文献来源:Travis Nemkov, Daniel Stephenson, Christopher Erickson, et al. Regulation of kynurenine metabolism by blood donor genetics and biology impacts red cell hemolysis in vitro and in vivo. Blood. 2024;143(5):456-472. doi: 10.1182/blood.2023022052.
抑制RHOA活性可维持长期低温保存血小板的存活和止血功能
编译:蔡仲仁,王浩阳 审校:伍昌林
1研究背景
造血干细胞移植受者和肿瘤患者需要持续进行血小板输注以降低出血风险,低温保存的血小板在数小时后会被宿主循环清除。因此,传统低温保存血小板被认为不适用于骨髓衰竭性血小板减少症患者的出血预防治疗,这类患者需将供体血小板在室温下储存不超过5天以防止细菌污染。然而,室温保存血小板的短保期限限制了输血可用性,且无法消除脓毒症风险。化学诱导病原体灭活虽获批准,但同样无法消除与脓毒症相关的细菌污染风险,且会降低血小板制品效力,导致为达到同等血小板计数增幅而增加输血需求。低温保存可显著减少微生物的增殖,延长储存周期,从而缓解当前地方层面血小板成分库存季节性短缺问题。
2主要结果
2.1低温保存诱导血小板活化和膜GPIbα/GPVI丢失
将儿童规格的抗凝血100%人血小板制品储存在透气聚烯烃袋中并持续振荡冷藏时,会引发血小板活化现象。通过检测活化GpIIb/IIIa表达水平、膜P-选择素及膜磷脂酰丝氨酸残基暴露(PS+)等指标,在储存第7天和第14天观察到显著变化。与RT血小板相比,低温保存血小板不仅丧失了膜GPIbα和GPVI活性,还伴随整体微粒和PS+微粒脱落量增加,同时肝细胞介导的血小板吞噬作用增强。该数据与既往研究结论一致:长期储存的血小板在阿司匹林治疗个体中会出现止血活性下降现象,且体内存活率降低。
2.2低温保存血小板早期激活RHOA和RAC1,而非CDC42
研究者发现小鼠RHOA和RAC1在低温保存条件下激活(图1A),小鼠血小板中的RAC最早在低温保存15分钟后被激活,随后RHOA在30分钟后被激活。本研究团队已开发出合理设计的、可穿透细胞的RHOA和RAC药理学抑制剂,这些抑制剂能有效阻止血小板中这两种蛋白的激活,且不会产生血小板毒性。RHOA抑制剂R-G04和RAC抑制剂NSC23766(NSC)在血小板的特异性浓度分别为75μM和50μM,显示出显著效果。类似地,CDC42抑制剂CASIN在血小板的特异性浓度为10μM时同样有效。通过分析三种抑制剂(R-G04、NSC和CASIN)的联合使用是否能通过阻止小鼠冷储存血小板的清除而产生有益效果。实验数据显示,抑制这三种Rho亚家族GTP酶可使未处理的受体(如RT储存血小板)保持存活状态(图1B-D)。
2.3药物性靶向RHOA足以预防冷诱导的血小板清除并维持止血活性
当存在载体或CDC42抑制剂CASIN时,人血小板在低温储存状态下被快速清除(曲线下面积AUC较常温储存组降低约57%);而使用RHOA抑制剂R-G04单独或与RAC抑制剂NSC联用时,血小板在体内的存活率与对照组常温储存组相当。使用RAC抑制剂NSC进行低温储存仅能部分延缓冷诱导的血小板清除现象(图1G-H)。体外实验:在储存第7天结束时对人血小板进行洗涤处理后,分别与活化的THP-1分化巨噬细胞共培养,观察其是否受抑制剂影响。与体内输血模型类似,使用RHOA抑制剂R-G04可有效阻断冷储存血小板的吞噬作用,而CDC42抑制剂CASIN则无此效果。在RAC抑制剂NSC存在的情况下,仅能部分抑制冷储存血小板的吞噬作用(图1I)。
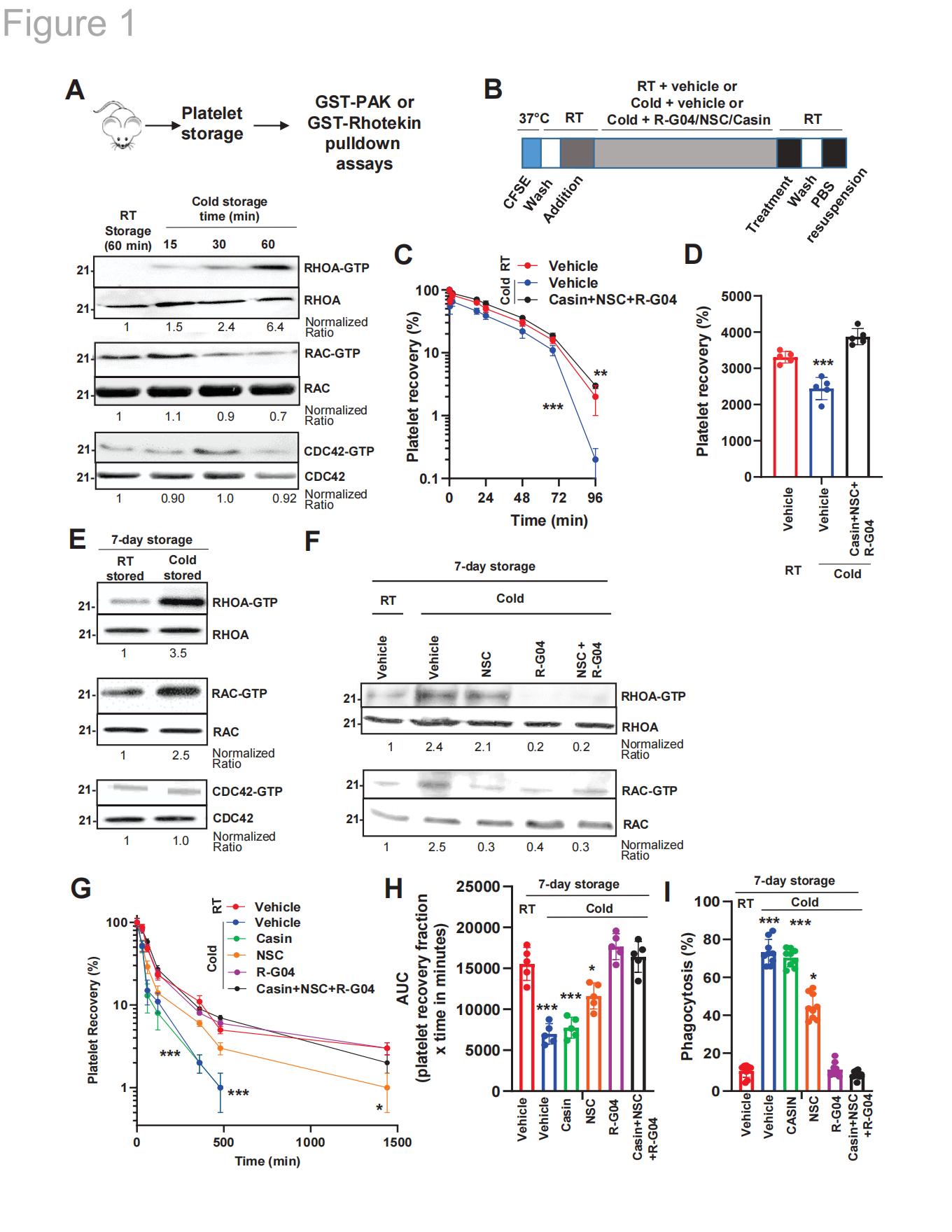
图1.小鼠低温保存血小板在冷藏后呈现RHOA激活现象,且同系输注后存活率降低;RHOA抑制剂可抑制低温保存人血小板的RHOA活性
2.4 RHOA的基因缺失可防止低温保存诱导的血小板清除
通过向Mx1-Cre;RhoAflox/flox小鼠(及对照)腹腔注射两剂聚肌胞苷酸(poly I:C)(剂量为10 mg/Kg/隔日),成功诱导血小板RHOA缺陷。在最后1次注射后的第3天,Mx1-Cre;RhoAflox/flox小鼠的血小板计数减少约50%,血小板RHOA表达量降低约90%(图2A)。小鼠RHOA缺陷型血小板在唾液酸、N-乙酰葡糖胺或半乳糖-N-乙酰葡糖胺相关凝集素的结合能力方面未见显著差异,即使经过冷冻保存后,其膜β3整合素(CD61)的表达水平仍保持稳定。血小板使用羧基荧光素琥珀酰亚胺酯(CFSE)进行标记。实验发现CFSE标记不干扰与低温保存相关糖基化结构相关的凝集素结合。将血小板调整至3×108(溶于200μL PBS),并输注至经辐照处理的血小板减少同源C57Bl/6小鼠体内。存活动力学分析表明,RhoAΔ/Δ血小板对低温保存完全不敏感,其体内寿命与RT保存血小板相当(图2B-C)。通过体外巨噬细胞介导的吞噬实验进一步证实,RHOA缺陷血小板对低温储存不敏感,且R-G04、NSC或其组合均未降低低温储存血小板的吞噬效率(图2D)。
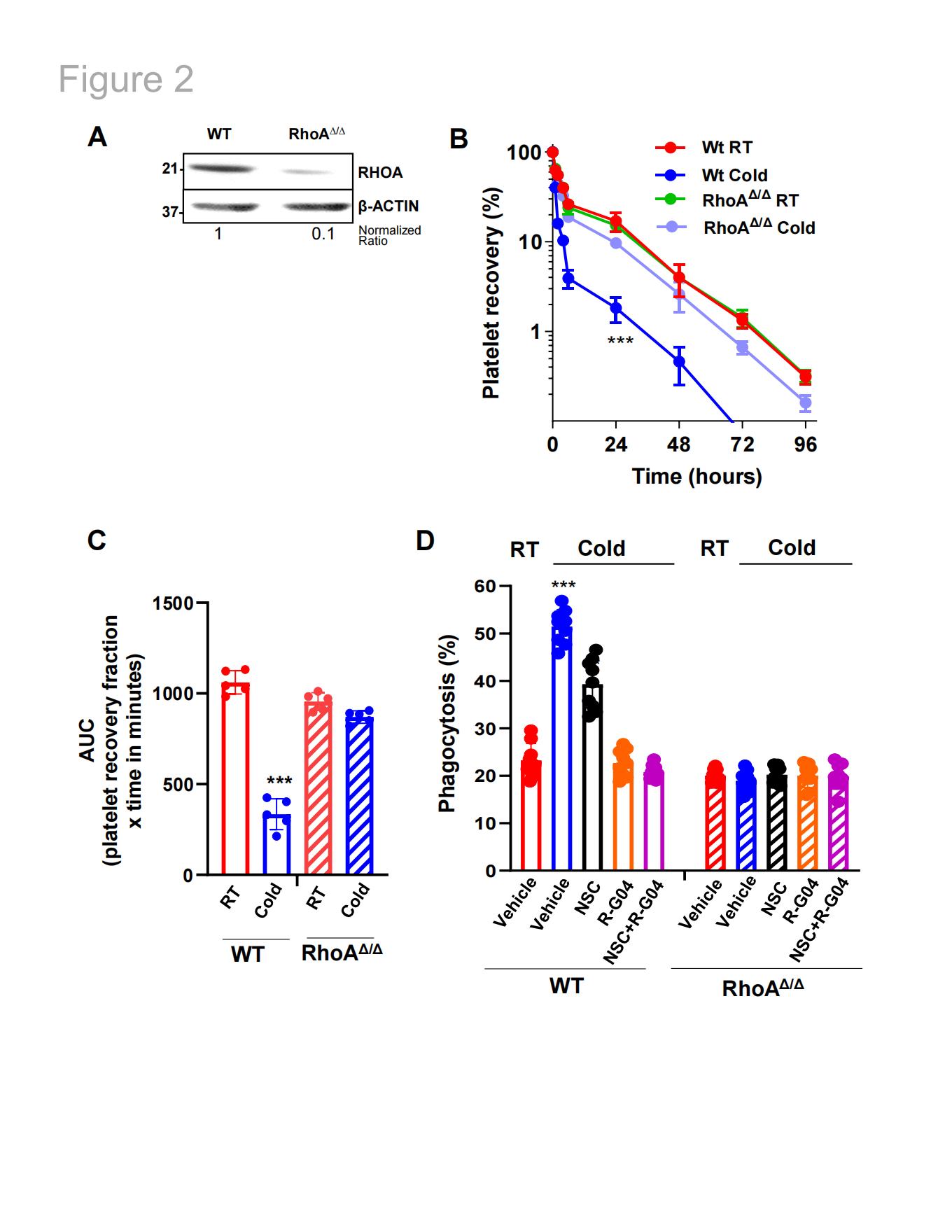
图2. RHOA缺失可预防血小板体内外清除
2.5 R-G04中的低温保存可以保持扩散,防止微粒脱落和肌动球蛋白激活
研究发现,经过7天低温保存处理的人血小板在固定化纤维蛋白原表面会形成微聚集体和直径小于1μm的血小板碎片。与未聚集、无微粒且表达肌动蛋白的血小板相比(图3B,溶剂处理组,低温处理血小板轮廓区域),这些血小板的铺展面积减少约70%,表现为应力纤维发育不全及皮层肌动蛋白聚合丧失(图3B-C)。与之相反,经R-G04处理的冷冻保存血小板在保留其铺展面积和形态方面表现优异,其水平与常规室温保存的对照组相当(图3B-C)。透射电镜分析显示,R-G04的存在显著抑制了冷冻保存血小板的微粒形成和囊泡脱落现象(图3D-E)。研究者观察到R-G04冷冻保存的血小板开放管腔系统的扩张基本被阻断(图3F),而通过RHOA药物靶向治疗可完全阻止肌球蛋白轻链(MLC,Ser19磷酸化)的激活,仅靶向RAC则只能部分抑制该过程(图3G)。这些数据表明:R-G04处理足以预防冷冻保存损伤引发的主要细胞学和结构变化,同时也能阻断影响肌动球蛋白激活的外向内信号传导。
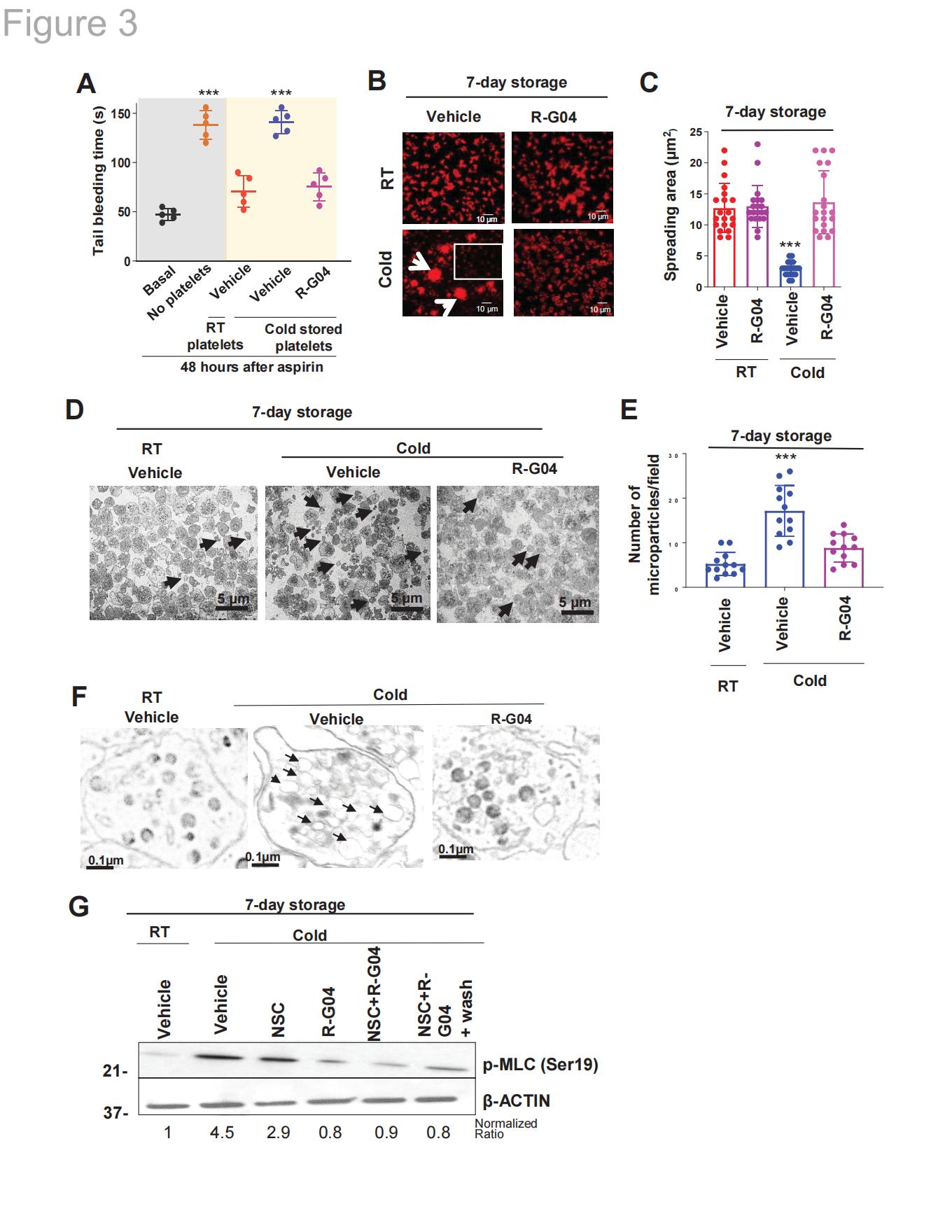
图3.R-G04介导的RHOA抑制具有可逆性,可恢复血小板体内凝血活性并预防低温保存相关的黏附缺陷、微粒形成及MLC激活
2.6靶向RHOA可预防Gplbα异常膜分布、膜脂筏形成及14-3-3ζ向低温保存人血小板脂筏中间体中的Gplbα募集
研究发现低温保存会诱导形成高表达GPIbα的膜区域(图4A),这种异常的GPIbα分布被R-G04所抑制,NSC23766则在较小程度上发挥作用,分析平面脂筏中的flotillin-1和洞状脂筏中的caveolin-1表达情况,发现冷藏保存的小鼠血小板(图4B)和保存7天的人类血小板(图4C-D)的膜中,富含flotilin-1和caveolin-1的脂筏。R-G04酚酞复现胆固醇耗竭剂甲基环糊精(MCD)的作用。相比之下,单独使用R-G04则能完全阻断冷冻保存对小鼠血小板(图4B)及储存3或7天的人类血小板(图4C-D)中平面脂筏和洞状脂筏生成的影响。作为RHOA活性下游多个效应因子之一的强效特异性ROCK抑制剂法舒地尔(Fasudil),既未表现出与R-G04相似的表型效应,也未能阻止冷储存血小板的吞噬作用。R-G04还能有效阻断冷储存引发的14-3-3ζ向Gplbα的募集过程,这种现象最终被诱导至脂筏中间体。
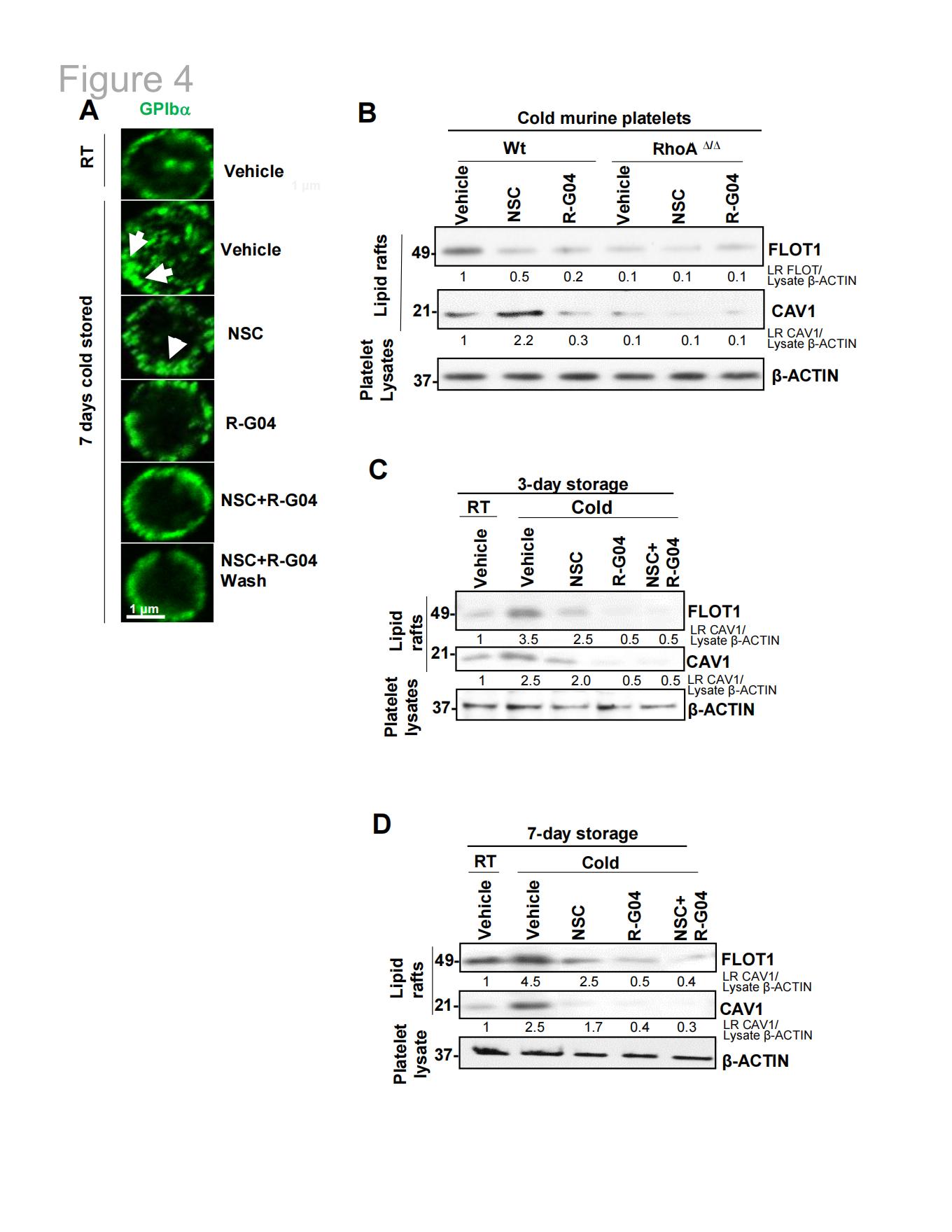
图4.血小板低温保存诱导GPIbα异常膜分布及脂筏形成,RHOA抑制可阻断此效应
2.7靶向RHOA可以完全防止相关糖基化酶在低温保存诱导下的错位
人血小板经冷藏处理后,GalNAc-T1/T3会形成膜簇状聚集(图5A-B)。研究发现,在冷藏过程中若存在R-G04或部分NSC成分,可有效抑制GalNAc-T1/T3的聚集现象(图5A-B)。洗涤步骤对未洗涤血小板中已识别的效应未产生显著影响。通过分析低温保存血小板膜脂筏提取物中UDP-半乳糖基转移酶和唾液酸转移酶的活性(这两种酶负责生成易受唾液酸酶作用的糖基结构),进一步证实这些关键糖基转移酶会转位至血小板膜微区(图5C-D)。R-G04和部分NSC可防止这些糖基转移酶活性的错误定位(图5C-D)。

图5. 抑制RHOA/RAC1可预防糖基化酶在低温诱导下错位定位
2.8 R-G04对映体S-G04的效力更强,可防止低温保存14天的人血小板清除
在低至10 μM浓度S-G04存在下,长期低温保存血小板可模拟75 μM R-G04的效应,足以防止血小板活化。S-G04能以与小鼠RHOA缺陷血小板相似的方式,有效阻止分化后THP-1巨噬细胞(图5E)或HepG2肝细胞对7天低温保存血小板的吞噬作用。在10μM S-G04存在下长期低温保存的人类血小板,在7或10天储存后维持输注血小板体内凝血活性的能力,与75μM R-G04相比表现相当(图5F-G)。经S-G04(10 μM)或R-G04(75 μM)处理的低温保存血小板能够有效防止异种输血后因冷冻保存导致的血小板清除现象(图6A)。该效果在储存7天、10天、以及14天(图6D-E)的血小板中均得到验证,与在止血功能模型中观察到的出血时间校正活性结果一致(图5F-G)。
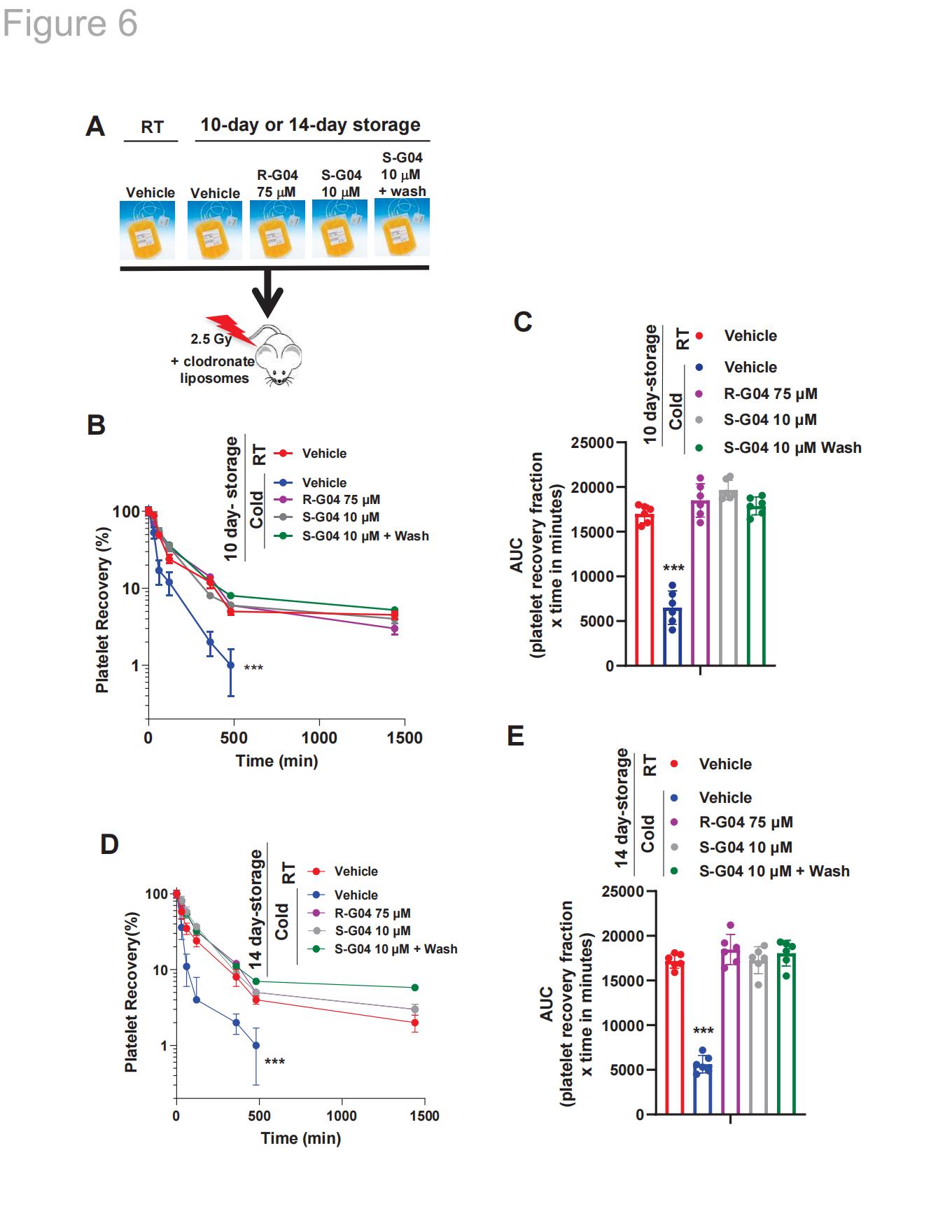
图6. S-G04作为R-G04的对映异构体,在临床前模型中表现出更强的效力,能够将低温保存人血小板的体内存活时间延长至10天
2.9 宿主AMR抑制和巨噬细胞耗竭的结合复制了R-G04和S-G04对维持人血小板存活的影响
与巨噬细胞耗竭效果类似(图6D-E),单独使用AMR抑制剂(图7A-C)无法阻止血小板清除,而预先储存的血小板在R-G04或SG04存在时则能有效阻断这一过程(图7A-C)。通过洗涤去除S-G04,其阻止血小板清除的作用并未改变(图7A-C)。只有当巨噬细胞同时耗竭且体内AMR被抑制时,长期低温保存的人类血小板在未添加R-G04或S-G04的环境中存活率才会变化(图7D-F)。这些血小板的存活水平与添加或未添加洗涤步骤的R-G04/S-G04处理组相比并无显著差异(图7D-E与图7B-C)。
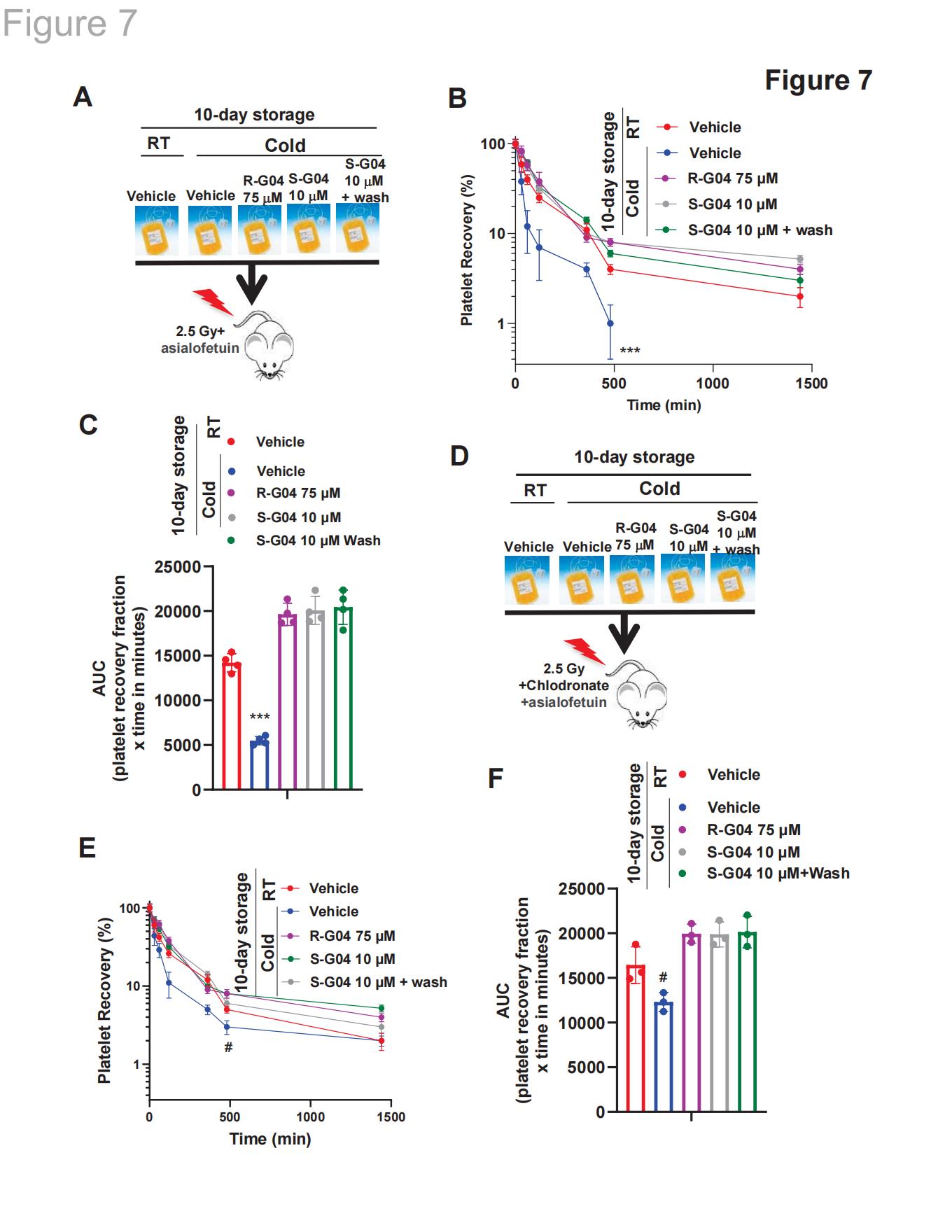
图7. 联合阻断巨噬细胞和肝细胞介导的清除作用重现了R-/S-G04对低温保存血小板中血小板清除的影响表型
3主要结论
1) 本研究揭示RHOA激活在血小板短期与长期低温保存损伤中的关键作用,低温保存会诱导RAC1和RHOA的活化,而RHOA基因缺失会导致血小板对低温保存诱导清除失去敏感性,RHOA是导致低温保存血小板清除过程中损伤的关键分子开关。
2) 通过使用R-G04或S-G04抑制RHOA,为预防低温保存引发的血小板储存损伤、避免输注时的清除风险、防止细菌污染,并将保存期限延长至储存后第2周内提供了有效解决方案。
文献来源:Hegde S, Akbar H, Wellendorf AM,et al. Inhibition of RHOA activity preserves the survival and hemostasis function of long-term cold-stored platelets.Blood. 2024;144(16):1732-1746. doi: 10.1182/blood.2023021453.
全文审校:李志强 潘 健
